Коацервация и связанная вода |
||
 |
 |
|
Оглавление
|
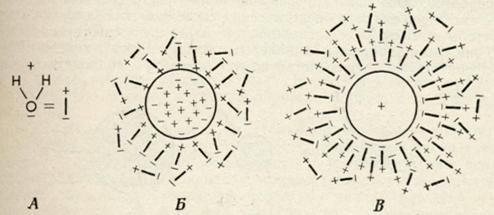
Коацервация и связанная водаКоллоидные системы обладают еще одним свойством, которое следует рассмотреть, прежде чем закончить обсуждение природы цитоплазматического матрикса. Это любопытное свойство, обусловленное действием поверхностных сил, известно как способность к образованию коацерватов. Необходимо вспомнить, что коллоидные частицы обладают двумя важными характеристиками — определенным размером и зарядом. Такие частицы будут ориентировать определенным образом окружающие их молекулы воды, образуя вокруг себя гидратную оболочку. Как показано на Рис. 1, непосредственно у поверхности частицы упорядоченность расположения молекул воды максимальна и постепенно убывает с расстоянием. Это означает, что в гомогенном коллоидном растворе будут присутствовать молекулы воды, весьма различные в структурном отношении: от молекул высокоструктурированной (связанной) воды и до молекул, ориентация которых столь же хаотична, как и в чистом растворителе. Постепенное уменьшение упорядоченности в расположении молекул воды вокруг коллоидной частицы приводит к исчезновению четкой границы, отделяющей ее от растворителя. Частицы при этом как бы заключены в своеобразный «матрикс», образо¬ванный молекулами растворителя. В наиболее простом случае, то есть в гомогенном коллоидном растворе, стабильность системы поддерживается благодаря влиянию двух факторов: отталкивания одноименно заряженных частиц и отделения их друг от друга сильно выраженными гидратными оболочками. Преципитация частиц в таких системах весьма маловероятна, и поэтому система в целом стабильна. Высказанные выше соображения можно использовать и для описания более сложных систем, в которых присутствуют частицы разноименного заряда, поскольку и в этом случае гидратные оболочки будут препятствовать агрегации частиц. В этих условиях устанавливается очень тонкое равновесие между силами притяжения разноименных зарядов, стремящихся сблизить коллоидные частицы, и силами отталкивания, то есть барьером который образуют их гидратные оболочки, препятствующие этому сближению. Система будет находиться в относительно устойчивом равновесии до тех пор, пока гидратные оболочки будут развиты в достаточной мере, чтобы выполнять функцию своеобразного буферного устройства, разделяющего частицы.
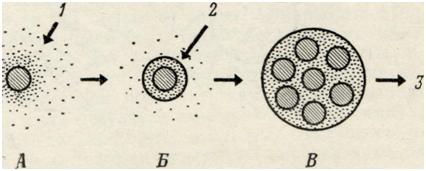
Введение в коллоидный раствор веществ, нейтрализующих заряд коллоидных частиц (ионов противоположного знака) или конкурирующих с ними за молекулы гидратационной воды (дегидратирующих агентов), нарушит строение гидратных оболочек. В связи с этим отчетливо выявится различие в свойствах свободной воды и воды, связанной коллоидными частицами. Говоря иными словами, коллоидные частицы при этом «сбросят» часть молекул воды, входящих в состав их гидратных оболочек, что приведет к внезапному образованию четкой границы, отделяющей эти частицы от общего объема коллоидного раствора. Размеры образующегося при этом гидратного комплекса определяются толщиной водной оболочки, которая все еще остается связанной коллоид¬ными частицами (Рис. 2). |
|||||||||||||
 |
 |
 |
Рис. 2 | ||
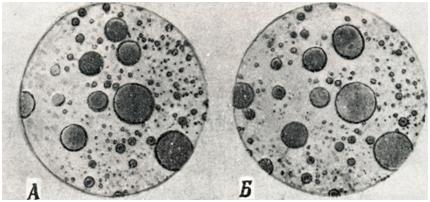
В описываемых нами условиях силы взаимного отталкивания одноименно заряженных частиц могут быть преодолены силами, стремящимися уменьшить поверхностную энергию системы, и частицы, сближаясь, будут образовывать агрегаты. Однако остаточные, хотя и сильно редуцированные, гидратные оболочки частиц все еще будут разделять их и препятствовать полному слиянию. Если указанный процесс будет продолжаться достаточно долго, то наступит флоккуляция и образование коацерватных капель, имеющих макроскопические размеры (фото I). Большая часть растворителя при этом удаляется, а образующиеся коацерватные капли представляют собой концентрированный раствор коллоида. Первоначально стабильная коллоидная система оказывается при этом разделенной на две четко различающиеся фазы, содержащие агрегаты коллоидных частиц различной концентрации.
|
 |
|
Фото I | ||
Следует иметь в виду, что одним из способов получения коацерватов является нейтрализация заряда частиц гомогенной коллоидной системы. При постепенной нейтрализации зарядов водная оболочка частиц становится менее компактной, а ее поверхность увеличивается. Поскольку заряд частиц при этом сильно уменьшается, они могут приблизиться друг к другу на достаточно близкие расстояния. С другой стороны, в том случае, когда коллоидный раствор содержит разнообразные частицы, отличающиеся как знаком, так и величиной заряда, потери гидратной оболочки бывает достаточно для образования сложных коацерватов. Этот последний случай более полно воспроизводит ситуацию, имеющую место в биологических системах, поскольку маловероятно, чтобы при большом разнообразии молекул, присутствующих в клетках, они были заряжены одинаковым образом. Хотя гидратные оболочки препятствуют непосредственному контакту коллоидных частиц, в некоторых случаях между ними может происходить образование межмолекулярных мостиков, что приводит к формированию геля. Однако этот способ образования геля не является типичным, и в большинстве случаев мы должны рассматривать гель и коацерваты как два различных состояния коллоидной системы. При коацервации образуются агрегаты, обладающие выраженной поверхностью. Именно этот аспект процесса коацервации был подвергнут всестороннему изучению, поскольку он важен для понимания происхождения клеточной структуры (в период возникновения жизни). В настоящем изложении процесс коацервации может быть использован в качестве модели для объяснения процессов, связанных с образованием некоторых видов клеточных вакуолей.
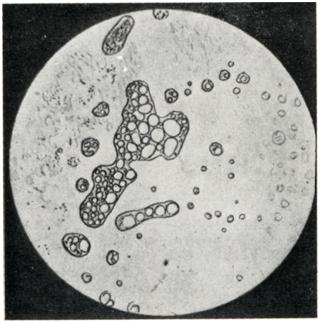
Вакуоли клеток (за исключением сократительной вакуоли, обладающей выраженной структурой) представляют собой преимущественно места отложения внеклеточных веществ, которые по большей части являются конечными продуктами клеточного обмена. Вакуоль обычно содержит 95—98% воды. Хотя в вакуоли наряду с большим количеством низкомолекулярных конечных продуктов метаболизма и солями присутствуют белки и иные высокомолекулярные соединения, ее содержимое редко находится в состоянии геля. Вакуоль растительной клетки может занимать большую часть объема клетки. В клетках животных чаще встречаются не одна большая, а много маленьких вакуолей. По существу то же самое мы наблюдаем и в молодых растительных клетках, где вакуоль либо вообще отсутствует, либо наблюдается множество мелких вакуолей. При изучении развития растительной клетки становится очевидным, что центральная вакуоль возникает благодаря слиянию ряда вакуолей меньшего размера, образующихся в цитоплазматическом матриксе. Можно показать, что жидкость, заполняющая эти вакуоли, есть результат коацервации, вызванной в свою очередь агрегацией большого числа коллоидных молекул цитоплазматического матрикса. Этот процесс аналогичен уже рассмотренному выше процессу «сбрасывания» гидратных оболочек благодаря нейтрализации зарядов молекул, что, как можно ожидать, происходит в такой сложной среде, какой является цитоплазматический матрикс клетки. Такая нейтрализация, заканчивающая рост и развитие клетки, вызывает частичную потерю связанной воды. Однако эксудат в этом случае выделяется внутрь плазматической мембраны в виде маленьких капелек воды, называемых «капельными вакуолями». Аналогичный процесс, как это можно видеть на фото II, может наблюдаться и в неживых коацерватных системах.
|
 |
|
Фото II | ||
В приведенном выше примере растворитель выделился внутрь коацерватной капли, вследствие чего капельки растворителя, накапливаясь в этой капле, образовали ряд вакуолей. Аналогия между процессами вакуолизации, происходящими в коацерватных каплях, и развитием вакуолей в клетках настолько очевидна, что ее невозможно игнорировать. С другой стороны, попытка распространить эту аналогию на клетки, обладающие высокой структурной организацией, приводит к ряду затруднений.
Существование тонопласта (вакуолярной мембраны растительной клетки) позволяет думать, что явление коацервации объясняет только отдельные стороны процесса образования вакуоли. Учитывая строение липопротеидной мембраны вакуоли, трудно предположить ее самопроизвольное образование вокруг коацерватной капли. Следует скорее предположить, что эта мембрана является структурой, производной от эндоплазматической сети клетки. Вероятнее всего, вакуолярная мембрана (в отношении как своего строения, так и выполняемой ею функции) во многом подобна цитоплазматической мембране окружающей клетку. Поэтому можно рассматривать содержимое вакуоли таким образом, как если бы оно находилось вне клетки. Во всяком случае, в рамках настоящего изложения наиболее существенным является то обстоятельство, что образование жидкости в клетке мы можем теперь связывать с коллоидными взаимодействиями и, таким образом, рассматривать этот процесс как закономерное следствие, вытекающее из свойств гидрофильных коллоидов цитоплазматического матрикса клетки. После образования вакуоли обмен веществ через ее мембрану подчиняется тем же закономерностям, которые определяют обмен клетки в целом.
Из проведенного обсуждения становится очевидным присутствие в цитоплазме двух «типов» воды: связанной (прочно удерживаемой коллоидными частицами) и свободной. Предполагается, что неструктурированная (или свободная) вода постоянно выделяется в вакуоли и что часть присутствующей в цитоплазме воды «связана» белками цитоплазмы. В сильно обезвоженных клетках, таких, например, как споры или семена, свободная вода совершенно отсутствует. Все гидрофильные гели после высушивания жадно поглощают воду. Тенденция коллоидных частиц к образованию гидратных оболочек столь велика, что гидрофильные гели набухают с большой силой. Действительно, развиваемое при этом давление в некоторых случаях достаточно для того, чтобы разрушить стенки стального сосуда.
Молекулы воды, входящие в состав коллоида, образуют весьма упорядоченную структуру. Хотя эта структура не является кристаллической, некоторые ее свойства близки к свойствам льда. При набухании высушенных коллоидов также происходит упорядочивание молекул воды и образование структур, напоминающих ледяные кристаллы. Благодаря этому довольно большое количество воды в клетках оказывается «структурированным», причем характер образующейся структуры определяется природой и строением коллоида. Многие клетки переносят замораживание без повреждений (важно, чтобы при этом не происходило образования больших кристаллов льда). Поэтому можно думать, что вода в этих клетках еще до замораживания находилась в высокоупорядоченном состоянии. Замораживание и оттаивание почти совершенно не изменяют жизнеспособности хорошо просушенных спор и семян. Вероятно, вся вода, содержащаяся в таких клетках, является связанной и характеризуется полукристаллической структурой.
Способность гидрофильных коллоидов удерживать воду важна для жизни вообще. Вся метаболическая активность клеток зависит от наличия водной среды; поэтому очевидно, что структура, которая позволяет клетке поддерживать определенную форму и вместе с тем придает ей высокую устойчивость к высушиванию, обеспечивает ей наибольшие шансы на выживание. Способность клеток переходить из метаболически активного состояния (гидратированный коллоид) в функционально неактивное, но стабильное состояние (спору) возможна лишь потому, что вода в этих объектах может существовать как в свободном, так и в связанном состоянии. Количество связанной воды, содержащееся в коллоидах, оказывается достаточным для поддержания внутренней организации споры, так что при достаточной гидратации она продолжает свой рост. В то же время удаление слабо связанных гидратных оболочек тотчас же вызывает приостановку обмена веществ и переводит клетку в покоящееся состояние. Интересной областью исследования является изучение вопроса о том, почему некоторые клетки легко переносят такие изменения своего состояния, в то время как другие при этом погибают. Причины, определяющие существование этих различий в свойствах клеток, в настоящее время еще совершенно неизвестны.
|
Иост Х. |

|
⇐ Перейти на главную страницу сайта |
⇑ Вернуться в начало страницы ⇑ |
Библиотека Ordo Deus ⇒ |
⇐ Вода свободная и связанная |
⇓ Каталог систематический ⇓ |
Зерновые корма ⇒ |
|
Внимание! Вы находитесь в библиотеке «Ordo Deus». Все книги в электронном варианте, содержащиеся в библиотеке «Ordo Deus», принадлежат их законным владельцам (авторам, переводчикам, издательствам). Все книги и статьи взяты из открытых источников и размещаются здесь только для чтения. |
|
Библиотека «Ordo Deus» не преследует никакой коммерческой выгоды. |
|
Все авторские права сохраняются за правообладателями. Если Вы являетесь автором данного документа и хотите дополнить его или изменить, уточнить реквизиты автора, опубликовать другие документы или возможно вы не желаете, чтобы какой-то из ваших материалов находился в библиотеке, пожалуйста, свяжитесь с нами по e-mail: |
Вас категорически не устраивает перспектива безвозвратно исчезнуть из этого мира? Вы желаете прожить ещё одну жизнь? Начать всё заново? Исправить ошибки этой жизни? Осуществить несбывшиеся мечты? Перейдите по ссылке: «главная страница».
|
© Ordo Deus, 2010. При копировании ссылка на сайт http://www.ordodeus.ru обязательна. |